
Scott Sherr, MD | Lucía González Forte, PhD
Submitted for publication [Article] to the SSRP Institute
Abstract
Methylene blue (MB) is a century-old compound whose renewed relevance in clinical medicine reflects a clearer understanding of its role in mitochondrial function, redox biology, and neuroinflammatory signaling. At therapeutic oral doses, MB acts as a catalytic electron shuttle within the mitochondrial respiratory chain, sustaining ATP production under conditions of dysfunction while limiting pathological reactive oxygen species generation. This bioenergetic effect extends to activation of the Nrf2/ARE antioxidant defense pathway and selective inhibition of inducible nitric oxide synthase — three mechanistically integrated actions that position MB as a multi-target compound rather than a conventional single-pathway drug. Clinical and preclinical evidence reflects this mechanism with meaningful strength: randomized data support adjunctive use in bipolar disorder, neuroimaging data confirm bioenergetic enhancement in healthy human subjects, and emerging evidence extends MB's redox profile into antimicrobial, antiviral, and post-viral fatigue applications. MB's effects are strongly dose-dependent, with benefits concentrated within a well-characterized low-dose oral window. This review summarizes the mechanistic foundation, key evidence, and clinical considerations for practitioners working in the cellular medicine space.
Introduction — A 150-Year-Old Molecule With a New Biological Identity

More than three billion people are currently living with a neurological condition, making neurological disorders the leading cause of illness and disability globally (GBD 2021 Nervous System Disorders Collaborators, 2024).
By 2050, dementia alone is projected to affect nearly 153 million people worldwide (Lin et al., 2025). Against this scale of unmet need, the pharmacological toolkit for chronic, progressive, and treatment-resistant neurological conditions has remained largely unchanged for decades — making the renewed scientific interest in a 150-year-old molecule not merely curious, but clinically significant.
Methylene blue was first synthesized in 1876 and entered medicine within a decade — initially as a biological stain, then in 1891 as the first fully synthetic drug used in clinical therapy, applied to treat malaria (Howland, 2016). By the 1930s, it had become the standard of care for methemoglobinemia, a condition in which hemoglobin loses its ability to carry oxygen, and it remains the only FDA-approved pharmacological intervention for that indication today (Seitkazina et al., 2022). For most of the twentieth century, its clinical identity was defined by these applications. The renewed interest of the past two decades represents something qualitatively different: the recognition that MB's redox chemistry has broader biological relevance than previously understood — specifically, that it participates directly in the cell's most foundational process: the production of energy itself.
What makes this renewed interest credible is not the molecule's age but what mechanistic research has revealed. MB preferentially accumulates in metabolically active tissues, crosses the blood-brain barrier readily, achieves oral bioavailability of approximately 72%, and reaches central nervous system concentrations substantially higher than systemic circulation within one hour of administration (Rojas et al., 2012; Walter-Sack et al., 2009). These pharmacological properties align precisely with the clinical contexts in which mitochondrial underperformance is implicated — cognitive decline, neuropsychiatric disorders, post-ischemic recovery, and increasingly, post-viral fatigue syndromes. The question is no longer whether MB has relevant biology. The question is how far that biology translates into improved clinical outcomes.
Mechanisms of Action — Three Convergent Pathways
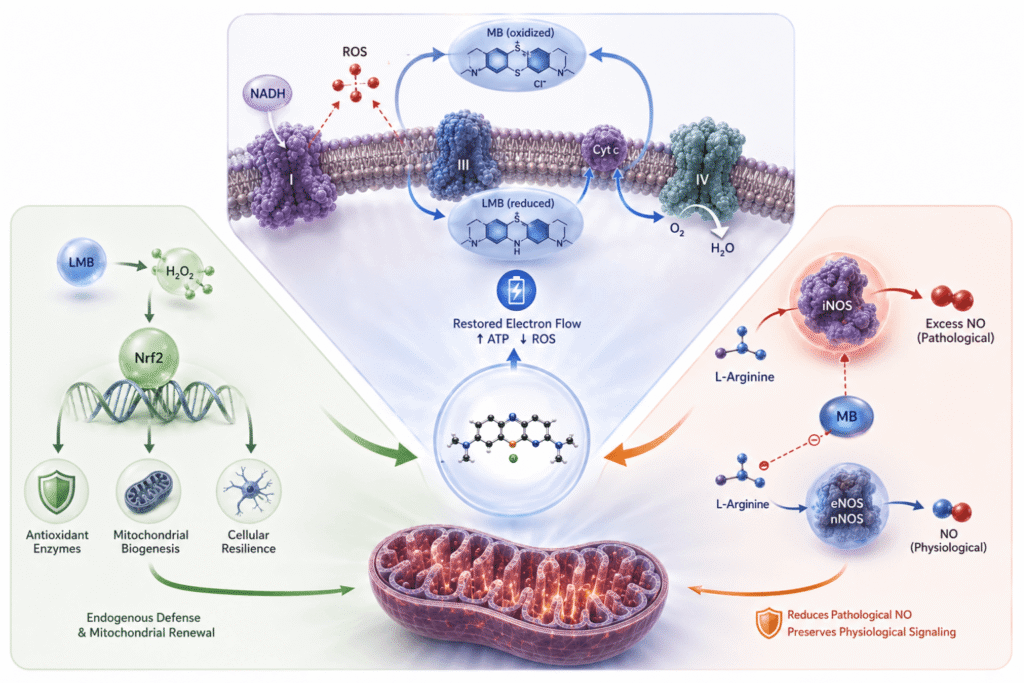
Methylene blue is pharmacologically unusual in that its primary site of action is not a receptor or enzyme in the conventional sense, but the mitochondrial electron transport chain itself. Three mechanistically integrated pathways account for the majority of its observed biological activity.
MB as a Mitochondrial Electron Shuttle
Under conditions of respiratory chain impairment — whether from hypoxia, neurotoxic injury, or the metabolic friction of aging — electrons leak prematurely from Complexes I and III, generating pathological reactive oxygen species (ROS) and impairing ATP synthesis (Wen et al., 2011). MB intervenes as a biological bypass circuit. In its oxidized form, it accepts electrons from NADH; once reduced to leucomethylene blue (LMB), it delivers electrons directly to cytochrome c upstream of Complex IV, partially restoring electron flow regardless of upstream impairment (Rojas et al., 2012; Gureev et al., 2022). Unlike conventional antioxidants consumed in a single reaction, MB is catalytically regenerated — functioning as a continuous redox cycler that stabilizes the bioenergetic engine rather than simply neutralizing individual ROS events.
This mechanism has been confirmed in the living human brain. In a randomized, double-blind, placebo-controlled fMRI trial, a single oral dose of MB enhanced neural activity during sustained attention and memory tasks, producing a 7% improvement in memory retrieval compared to placebo (Rodriguez et al., 2016). Rodent fMRI studies further demonstrated augmented cerebral oxygen consumption and blood flow under both normoxic and hypoxic conditions (Lin et al., 2012; Huang et al., 2013). The bioenergetic mechanisms characterized in animal models are measurably active in the living human brain.
Nrf2 Activation and Endogenous Antioxidant Defense
A secondary consequence of MB's redox cycle is the controlled generation of hydrogen peroxide (H₂O₂) as a byproduct of LMB re-oxidation (Tretter et al., 2014). At therapeutic doses, this H₂O₂ is modest and localized — functioning as a redox signaling molecule that activates the Nrf2/ARE pathway, the cell's central regulator of endogenous antioxidant defense. Nrf2 activation drives transcription of key antioxidant enzymes and, critically, promotes mitochondrial biogenesis — the generation of new mitochondria — with particular relevance for neurons and other high-demand tissues (Tucker et al., 2018; Samoylova et al., 2023). This self-amplifying response extends well beyond MB's immediate presence, and distinguishes it mechanistically from direct ROS scavengers, which have shown limited and inconsistent efficacy in neurological contexts precisely because they do not address upstream bioenergetic impairment.
Selective Modulation of Nitric Oxide Signaling
A third mechanism operates through MB's inhibition of inducible nitric oxide synthase (iNOS) and soluble guanylate cyclase (sGC). Constitutive nitric oxide production by endothelial and neuronal NOS isoforms is essential for vascular tone and synaptic plasticity. The clinical problem arises from iNOS, which is upregulated during inflammation and can generate nitric oxide at levels that drive tissue damage and neuroinflammatory cascades. MB's pharmacological relevance lies in its selective affinity for iNOS at low doses, while largely sparing eNOS and nNOS — attenuating pathological NO overproduction without disrupting constitutive signaling on which normal vascular and neuronal function depends (Gureev et al., 2022; Pluta et al., 2023).
Taken together, these three mechanisms — electron shuttle, Nrf2-mediated cellular defense, and iNOS-selective NO modulation — are hierarchically integrated: a core bioenergetic intervention, downstream adaptive antioxidant responses, and parallel modulation of inflammatory signaling through a single, catalytically regenerated molecule.
Evidence Base — Preclinical Coherence and Clinical Translation
Preclinical Record
The preclinical literature for MB is notable not simply for its breadth but for the consistency of findings across independent research groups, species, and model types. In a rat model of chronic cerebral hypoperfusion relevant to vascular dementia, MB attenuated learning and memory deficits while preserving cytochrome oxidase activity and reducing neurodegeneration in affected brain regions (Auchter et al., 2014; Auchter et al., 2020). In transgenic Alzheimer's models, MB reduces amyloid-beta accumulation by increasing proteasome activity and inhibits Caspase-6, reversing both cognitive decline and neuroinflammatory markers in aged mice (Medina et al., 2011; Zhou et al., 2019).
In ischemic stroke models, MB produces a significant reduction in infarct volume through preservation of the penumbra, stimulation of autophagy, inhibition of apoptosis, and promotion of post-ischemic neurogenesis (Shen et al., 2013; Jiang et al., 2015; Ahmed et al., 2016). Across traumatic brain injury models, a single intravenous dose administered within 30 minutes of injury reduced neuronal apoptosis, preserved blood-brain barrier integrity, improved ATP production, and attenuated neuroinflammation and depressive-like behavior — effects persisting up to 180 days with continued dosing (Watts et al., 2014; Shen et al., 2019; Fenn et al., 2015). A 2024 review in Reviews in the Neurosciences synthesized this converging evidence, positioning MB as a multi-indication neuroprotective agent whose therapeutic window is greatest when administered within hours of injury (Isaev et al., 2024).
Clinical Evidence
The strongest clinical evidence for MB is in bipolar disorder. In a randomized, double-blind, placebo-controlled crossover trial, MB at 195 mg/day significantly reduced residual depressive and anxiety symptoms without precipitating mania — a clinically important safety finding given the risk profile of most augmentation strategies in this population (Alda et al., 2017). The mechanism involves both mitochondrial bioenergetic support and MAO-A inhibition, which elevates serotonin, dopamine, and norepinephrine in a dose-dependent manner (Harvey et al., 2010; Alda, 2019). The neuroimaging evidence in healthy humans confirming a 7% memory improvement at low oral doses establishes that these mechanisms are active and clinically meaningful even in the absence of diagnosed pathology (Rodriguez et al., 2016).
In Alzheimer's disease, MB's potential has been investigated primarily through tau aggregation inhibition. Phase II trials of MB-derived compounds showed signals of cognitive benefit; the subsequent Phase III trial of LMTM — a chemically modified derivative — failed to demonstrate superiority over its low-dose comparator, a result complicated by the likelihood that both arms were biologically active at the doses used (Gauthier et al., 2016). This result should not be read as a straightforward negative for MB itself, as the preclinical evidence for MB's effects on amyloid clearance via proteasome activation and inhibition of neuronal self-destruction pathways operates through mechanisms distinct from and independent of tau aggregation (Medina et al., 2011).
Emerging Applications
Beyond established neuropsychiatric indications, MB's redox chemistry supports several emerging clinical applications.
In antimicrobial contexts, photoactivation of MB by red light (~660 nm) generates reactive oxygen species via both radical and singlet oxygen pathways, producing multi-target oxidative activity against microbial membranes with a mechanism independent of classical antibiotic resistance pathways. In vitro, MB-mediated antimicrobial photodynamic therapy has demonstrated broad-spectrum bactericidal activity against WHO global priority pathogens, including multidrug-resistant strains of Acinetobacter baumannii, Klebsiella pneumoniae, MRSA, and vancomycin-resistant Enterococcus, with no correlation observed between antibiotic resistance profiles and photodynamic sensitivity (Songsantiphap et al., 2022; Sabino et al., 2020). Early in vitro evidence also identifies MB among the most active agents against biofilm-forming Bartonella henselae and Borrelia burgdorferi persisters — organisms for which conventional antibiotic therapy has demonstrated persistent limitations (Zheng et al., 2020; Feng et al., 2015).
Long COVID represents one of the most mechanistically relevant emerging indications. At least 65 million people globally meet criteria for Long COVID, and converging clinical and laboratory evidence now implicates mitochondrial dysfunction as a central driver of its most prevalent symptoms — fatigue, cognitive impairment, and post-exertional malaise. Reduced mitochondrial enzyme activity and structural mitochondrial abnormalities have been documented in Long COVID patients (Appelman et al., 2024; Macnaughtan et al., 2025), and SARS-CoV-2 infection has been shown to directly suppress core mitochondrial gene expression and impair mitophagy (Guarnieri et al., 2023). As an electron shuttle capable of bypassing impaired respiratory chain complexes, an activator of Nrf2-driven antioxidant defense, and a selective iNOS inhibitor, MB's mechanistic profile addresses the precise bioenergetic failure implicated in post-viral pathophysiology (Molnar et al., 2024). Clinical trial data in this population remain absent, but the mechanistic alignment is among the strongest for any currently investigational agent.
Clinical Application and Safety Considerations

Dosing and Hormetic Dose-Response
Understanding how dose determines outcome is the prerequisite for both safe and effective MB use. Unlike most pharmacological agents, MB exhibits a well-characterized hormetic dose-response: the same electron-shuttling mechanism that enhances mitochondrial function at low doses generates excess oxidative stress at high doses, impairing the very processes it otherwise supports (Rojas et al., 2012; Tucker et al., 2018). This biphasic pattern is among the most consistently replicated findings in the MB literature and has a direct practical implication: dose selection determines whether MB acts as a bioenergetic support or a pro-oxidant stressor.
For neuromodulatory and bioenergetic applications, the evidence points to a therapeutic window of 0.2–4 mg/kg orally, with beneficial effects concentrated at the lower end of this range. The bipolar disorder trials used 195 mg/day (~2–3 mg/kg for an average adult), while doses as low as 15 mg/day showed measurable clinical activity compared to an inert placebo (Alda et al., 2017). Oral bioavailability is approximately 72%, and MB crosses the blood-brain barrier readily, reaching CNS concentrations substantially higher than systemic circulation within one hour of administration (Walter-Sack et al., 2009; Rojas et al., 2012). Sublingual or buccal administration bypasses first-pass hepatic metabolism and may support more consistent systemic exposure at lower nominal doses — a consideration with particular relevance given that target bioenergetic effects are achievable at the low end of the therapeutic window.
Formulation quality is a clinically relevant variable. Pharmaceutical-grade sourcing with independent purity verification is essential, as lower-grade preparations may contain trace heavy metals, including lead, mercury, cadmium, or arsenic, undermining both safety and clinical predictability.
Patient Selection Framework
The conditions where MB has the greatest current clinical relevance are those where mitochondrial dysfunction and neuroinflammation are mechanistically implicated: residual neuropsychiatric symptoms, early cognitive decline with vascular or metabolic contributors, post-ischemic recovery, TBI sequelae, and post-viral fatigue syndromes, including Long COVID. In bipolar disorder, the mechanistic rationale has been clinically validated; in the others, human trial data remain limited, but the biological case is strong.
A useful patient selection framework centers on three questions: Is mitochondrial dysfunction mechanistically plausible given the clinical picture? Is there evidence of neuroinflammatory burden? Do available conventional options adequately address the underlying biology? When the answer to the first two is yes, and the third is no, MB warrants consideration — not as a first-line therapy, but as a mechanism-aligned adjunct. Patient selection guided by biological plausibility rather than diagnostic label is likely to identify the populations that derive the greatest benefit.
Safety Profile and Clinical Precautions
MB has a well-characterized safety profile at low therapeutic doses. Adverse effects are generally mild and transient: blue-green discoloration of urine (an expected pharmacokinetic consequence), occasional nausea, headache, and dizziness (Seitkazina et al., 2022). Four specific conditions require active screening before initiation.
G6PD deficiency is an inherited enzymatic condition that may reduce the capacity to buffer the hydrogen peroxide generated by MB's redox cycle, with hemolysis as a potential consequence at high intravenous doses; the clinical picture at low oral doses is less clearly defined, with no reported cases of hemolysis at oral dosing levels in the literature (Youngster et al., 2010).
Serotonergic medications warrant individualized management due to MB's dose-dependent MAO-A inhibition. Documented cases of MB-induced serotonin syndrome are exclusively derived from high-dose intravenous administration (1–2 mg/kg IV); oral administration produces an AUC approximately 15-fold lower than equivalent intravenous doses, resulting in substantially lower peak plasma concentrations (Gillman, 2011; Zuschlag et al., 2018; Peter et al., 2000).
Hypertension and cardiovascular considerations arise from MB's inhibition of soluble guanylate cyclase, which increases systemic vascular resistance and mean arterial pressure in a dose-dependent manner (Kwok & Howes, 2006; Zhao et al., 2022). At low oral doses, the hemodynamic impact is substantially attenuated, but the mechanism remains active. Patients with poorly controlled hypertension, those taking nitrate-based vasodilators or PDE5 inhibitors, and those with significant cardiovascular disease warrant individualized risk assessment and active blood pressure monitoring during dose titration.
Renal impairment produces clinically meaningful increases in systemic MB exposure due to substantial renal excretion of MB and its metabolites. Pharmacokinetic data from the FDA-approved PROVAYBLUE label demonstrate AUC increases of 52%, 116%, and 192% in mild, moderate, and severe renal impairment, respectively, with the active metabolite Azure B accumulating even more steeply — 339% in severe impairment (US Food and Drug Administration, 2024). Therapy should be avoided in severe renal impairment and used with caution, with extended monitoring, in mild to moderate dysfunction.
Research Priorities
The most actionable research priorities for MB are: adequately powered randomized trials in ischemic stroke recovery, vascular cognitive impairment, traumatic brain injury, and Long COVID; dose-finding studies in each population given the clinical consequences of the hormetic dose-response; and clinical translation of the antimicrobial evidence in biofilm-forming and intracellular pathogens. What MB brings to each of these priorities is something increasingly rare: a deeply understood mechanism, a safety record spanning 150 years of clinical use, and oral bioavailability that makes clinical trial design straightforward.
SSRP Disclaimer:
The views, interpretations, and conclusions expressed are those of the author(s); publication does not imply endorsement by the SSRP Institute. SSRP editorial review confirms that Community Articles submissions meet SSRP's publication standards: grounded in peer-reviewed evidence, oriented toward disease-state mechanisms and clinical application, and free of product promotion. Editorial review does not constitute independent verification of cited research or clinical endorsement of the described applications. Publication of Community Article submissions are intended for education and informational purposes only, and should not be considered medical advice.
Conflict(s) of Interest:
Dr. Scott Sherr serves as the Chief Medical and Health Education Officer for Troscriptions, a company that develops and markets methylene blue-containing products. As part of this role, he contributes to product education, scientific communication, and clinical guidance. This relationship is disclosed in the interest of transparency. The scientific interpretations and conclusions presented in this article are based on the available published literature and current evidence.
References
Alda, M. (2019). Methylene blue in the treatment of neuropsychiatric disorders. CNS Drugs, 33(8), 719–725. https://doi.org/10.1007/s40263-019-00641-3
Alda, M., McKinnon, M., Blagdon, R., Garnham, J., MacLellan, S., O’Donovan, C., Hajek, T., Nair, C., Dursun, S., & MacQueen, G. (2017). Methylene blue treatment for residual symptoms of bipolar disorder: Randomised crossover study. British Journal of Psychiatry, 210(1), 54–60. https://doi.org/10.1192/bjp.bp.115.173930
Appelman, B., Charlton, B. T., Goulding, R. P., Kerkhoff, T. J., Breedveld, E. A., Noort, W., Offringa, C., Bloemers, F. W., van Weeghel, M., Schomakers, B. V., Coelho, P., Posthuma, J. J., Aronica, E., Wiersinga, W. J., van Vugt, M., & Wüst, R. C. I. (2024). Muscle abnormalities worsen after post-exertional malaise in long COVID. Nature Communications, 15(1), 17. https://doi.org/10.1038/s41467-023-44432-3
Auchter, A., Williams, J., Barksdale, B., Monfils, M. H., & Gonzalez-Lima, F. (2014). Therapeutic benefits of methylene blue on cognitive impairment during chronic cerebral hypoperfusion. Journal of Alzheimer's Disease, 42(S4), S525–S535. https://doi.org/10.3233/JAD-141527
Auchter, A., Barrett, D. W., Monfils, M. H., & Gonzalez-Lima, F. (2020). Methylene blue preserves cytochrome oxidase activity and prevents neurodegeneration and memory impairment in rats with chronic cerebral hypoperfusion. Frontiers in Cellular Neuroscience, 14, 130. https://doi.org/10.3389/fncel.2020.00130
Fenn, A. M., Skendelas, J. P., Moussa, D. N., Muccigrosso, M. M., Popovich, P. G., Lifshitz, J., Eiferman, D. S., & Godbout, J. P. (2015). Methylene blue attenuates traumatic brain injury-associated neuroinflammation and acute depressive-like behavior in mice. Journal of Neurotrauma, 32(2), 127–138. https://doi.org/10.1089/neu.2014.3514
Feng, J., Weitner, M., Shi, W., Zhang, S., Sullivan, D., & Zhang, Y. (2015). Identification of additional anti-persister activity against Borrelia burgdorferi from an FDA drug library. Antibiotics, 4(3), 397–410. https://doi.org/10.3390/antibiotics4030397
Gauthier, S., Feldman, H. H., Schneider, L. S., Wilcock, G. K., Frisoni, G. B., Hardlund, J. H., Moebius, H. J., Bentham, P., Kook, K. A., Wischik, D. J., Schelter, B. O., Davis, C. S., Staff, R. T., Bracoud, L., Shamsi, K., Storey, J. M. D., Harrington, C. R., & Wischik, C. M. (2016). Efficacy and safety of tau-aggregation inhibitor therapy in patients with mild or moderate Alzheimer's disease: A randomised, controlled, double-blind, parallel-arm, phase 3 trial. The Lancet, 388(10062), 2873–2884. https://doi.org/10.1016/S0140-6736(16)31275-2
GBD 2021 Nervous System Disorders Collaborators. (2024). Global, regional, and national burden of disorders affecting the nervous system, 1990–2021: A systematic analysis for the Global Burden of Disease Study 2021. Lancet Neurology, 23(4), 344–381. https://doi.org/10.1016/S1474-4422(24)00038-3
Gillman, P. K. (2011). CNS toxicity involving methylene blue: The exemplar for understanding and predicting drug interactions that precipitate serotonin toxicity. Journal of Psychopharmacology, 25(3), 429–436. https://doi.org/10.1177/0269881109359098
Guarnieri, J. W., Dybas, J. M., Fazelinia, H., Kim, M. S., Frere, J., Zhang, Y., Soto Albrecht, Y., Murdock, D. G., Angelin, A., Singh, L. N., Cope, H., Zaksas, V., Saravia-Butler, A., Meydan, C., Foox, J., Mozsary, C., Bram, Y., Kidane, Y., Priebe, W., . . . Wallace, D. C. (2023). Core mitochondrial genes are down-regulated during SARS-CoV-2 infection of rodent and human hosts. Science Translational Medicine, 15(708), eabq1533. https://doi.org/10.1126/scitranslmed.abq1533
Gureev, A. P., Sadovnikova, I. S., & Popov, V. N. (2022). Molecular mechanisms of the neuroprotective effect of methylene blue. Biochemistry (Moscow), 87(9), 940–956. https://doi.org/10.1134/S0006297922090073
Harvey, B. H., Duvenhage, I., Viljoen, F., Scheepers, N., Malan, S. F., Wegener, G., Brink, C. B., & Petzer, J. P. (2010). Role of monoamine oxidase, nitric oxide synthase and regional brain monoamines in the antidepressant-like effects of methylene blue and selected structural analogues. Biochemical Pharmacology, 80(10), 1580–1591. https://doi.org/10.1016/j.bcp.2010.07.037
Howland, R. H. (2016). Methylene blue: The long and winding road from stain to brain: Part 1. Journal of Psychosocial Nursing and Mental Health Services, 54(9), 21–24. https://doi.org/10.3928/02793695-20160818-01
Huang, S., Du, F., Shih, Y. Y., Shen, Q., Gonzalez-Lima, F., & Duong, T. Q. (2013). Methylene blue potentiates stimulus-evoked fMRI responses and cerebral oxygen consumption during normoxia and hypoxia. NeuroImage, 72, 237–242. https://doi.org/10.1016/j.neuroimage.2013.01.027
Isaev, N. K., Genrikhs, E. E., & Stelmashook, E. V. (2024). Methylene blue and its potential in the treatment of traumatic brain injury, brain ischemia, and Alzheimer's disease. Reviews in the Neurosciences, 35(5), 585–595. https://doi.org/10.1515/revneuro-2024-0007
Jiang, Z., Watts, L. T., Huang, S., Shen, Q., Rodriguez, P., Chen, C., Zhou, C., & Duong, T. Q. (2015). The effects of methylene blue on autophagy and apoptosis in MRI-defined normal tissue, ischemic penumbra and ischemic core. PLoS One, 10(6), e0131929. https://doi.org/10.1371/journal.pone.0131929
Kwok, E. S. H., & Howes, D. (2006). Use of methylene blue in sepsis: A systematic review. Journal of Intensive Care Medicine, 21(6), 359–363. https://doi.org/10.1177/0885066606290671
Lin, A. L., Poteet, E., Du, F., Gourav, R. C., Liu, R., Wen, Y., Bresnen, A., Huang, S., Fox, P. T., Yang, S. H., & Duong, T. Q. (2012). Methylene blue as a cerebral metabolic and hemodynamic enhancer. PLoS One, 7(10), e46585. https://doi.org/10.1371/journal.pone.0046585
Lin, C. Y., Zhai, Y. J., An, H. H., Zhou, J. X., & Zhao, Y. (2025). Global trends in prevalence, disability adjusted life years, and risk factors for early onset dementia from 1990 to 2021. Scientific Reports, 15, 13488. https://doi.org/10.1038/s41598-025-97404-6
Macnaughtan, J., Chau, K. Y., Brennan, E., Toffoli, M., Spinazzola, A., Hillman, T., Heightman, M., & Schapira, A. H. V. (2025). Mitochondrial function is impaired in long COVID patients. Annals of Medicine, 57(1), 2528167. https://doi.org/10.1080/07853890.2025.2528167
Medina, D. X., Caccamo, A., & Oddo, S. (2011). Methylene blue reduces Aβ levels and rescues early cognitive deficit by increasing proteasome activity. Brain Pathology, 21(2), 140–149. https://doi.org/10.1111/j.1750-3639.2010.00430.x
Molnar, T., Lehoczki, A., Fekete, M., Varnai, R., Zavori, L., Erdo-Bonyar, S., Simon, D., Berki, T., Csecsei, P., & Ezer, E. (2024). Mitochondrial dysfunction in long COVID: Mechanisms, consequences, and potential therapeutic approaches. GeroScience, 46(5), 5267–5286. https://doi.org/10.1007/s11357-024-01165-5
Peter, C., Hongwan, D., Küpfer, A., & Lauterburg, B. H. (2000). Pharmacokinetics and organ distribution of intravenous and oral methylene blue. European Journal of Clinical Pharmacology, 56(3), 247–250. https://doi.org/10.1007/s002280000124
Pluta, M. P., Putowski, Z., Czempik, P. F., & Krzych, L. J. (2023). Successful use of methylene blue in catecholamine-resistant septic shock: A case report and short literature review. International Journal of Molecular Sciences, 24(13), 10772. https://doi.org/10.3390/ijms241310772
Rodriguez, P., Zhou, W., Barrett, D. W., Altmeyer, W., Gutierrez, J. E., Li, J., Lancaster, J. L., Gonzalez-Lima, F., & Duong, T. Q. (2016). Multimodal randomized functional MR imaging of the effects of methylene blue in the human brain. Radiology, 281(2), 516–526. https://doi.org/10.1148/radiol.2016152893
Rojas, J. C., Bruchey, A. K., & Gonzalez-Lima, F. (2012). Neurometabolic mechanisms for memory enhancement and neuroprotection of methylene blue. Progress in Neurobiology, 96(1), 32–45. https://doi.org/10.1016/j.pneurobio.2011.10.007
Sabino, C. P., Wainwright, M., Ribeiro, M. S., Sellera, F. P., dos Anjos, C., da Silva Baptista, M., & Lincopan, N. (2020). Global priority multidrug-resistant pathogens do not resist photodynamic therapy. Journal of Photochemistry and Photobiology B: Biology, 208, 111893. https://doi.org/10.1016/j.jphotobiol.2020.111893
Samoylova, N. A., Gureev, A. P., & Popov, V. N. (2023). Methylene blue induces antioxidant defense and reparation of mitochondrial DNA in a Nrf2-dependent manner during cisplatin-induced renal toxicity. International Journal of Molecular Sciences, 24(7), 6118. https://doi.org/10.3390/ijms24076118
Seitkazina, A., Yang, J. K., & Kim, S. (2022). Clinical effectiveness and prospects of methylene blue: A systematic review. Precision and Future Medicine, 6(4), 193–208. https://doi.org/10.23838/pfm.2022.00079
Shen, J., Xin, W., Li, Q., Gao, Y., Yuan, L., & Zhang, J. (2019). Methylene blue reduces neuronal apoptosis and improves blood-brain barrier integrity after traumatic brain injury. Frontiers in Neurology, 10, 1133. https://doi.org/10.3389/fneur.2019.01133
Shen, Q., Du, F., Huang, S., Rodriguez, P., Watts, L. T., & Duong, T. Q. (2013). Neuroprotective efficacy of methylene blue in ischemic stroke: An MRI study. PLoS One, 8(11), e79833. https://doi.org/10.1371/journal.pone.0079833
Songsantiphap, C., Vanichanan, J., Chatsuwan, T., Asawanonda, P., & Boontaveeyuwat, E. (2022). Methylene blue–mediated antimicrobial photodynamic therapy against clinical isolates of extensively drug resistant Gram-negative bacteria causing nosocomial infections in Thailand, an in vitro study. Frontiers in Cellular and Infection Microbiology, 12, 929242. https://doi.org/10.3389/fcimb.2022.929242
Tretter, L., Horvath, G., Hölgyesi, A., Essek, F., & Adam-Vizi, V. (2014). Enhanced hydrogen peroxide generation accompanies the beneficial bioenergetic effects of methylene blue in isolated brain mitochondria. Free Radical Biology and Medicine, 77, 317–330. https://doi.org/10.1016/j.freeradbiomed.2014.09.024
Tucker, D., Lu, Y., & Zhang, Q. (2018). From mitochondrial function to neuroprotection — an emerging role for methylene blue. Molecular Neurobiology, 55(6), 5137–5153. https://doi.org/10.1007/s12035-017-0712-2
US Food and Drug Administration. (2024). PROVAYBLUE® (methylene blue) injection, USP: Prescribing information. https://www.accessdata.fda.gov/drugsatfda_docs/label/2024/204630s021lbl.pdf
Walter-Sack, I., Rengelshausen, J., Oberwittler, H., Burhenne, J., Mueller, O., Meissner, P., & Mikus, G. (2009). High absolute bioavailability of methylene blue given as an aqueous oral formulation. European Journal of Clinical Pharmacology, 65(2), 179–189. https://doi.org/10.1007/s00228-008-0563-x
Watts, L. T., Long, J. A., Chemello, J., Van Koughnet, S., Fernandez, A., Huang, S., Shen, Q., & Duong, T. Q. (2014). Methylene blue is neuroprotective against mild traumatic brain injury. Journal of Neurotrauma, 31(11), 1063–1071. https://doi.org/10.1089/neu.2013.3193
Wen, Y., Li, W., Poteet, E. C., Xie, L., Tan, C., Yan, L. J., Ju, X., Liu, R., Qian, H., Marvin, M. A., Goldberg, M. S., She, H., Mao, Z., Simpkins, J. W., & Yang, S. H. (2011). Alternative mitochondrial electron transfer as a novel strategy for neuroprotection. Journal of Biological Chemistry, 286(18), 16504–16515. https://doi.org/10.1074/jbc.M110.208447
Youngster, I., Arcavi, L., Schechmaster, R., Akayzen, Y., Popliski, H., Shimonov, J., Beig, S., & Berkovitch, M. (2010). Medications and glucose-6-phosphate dehydrogenase deficiency: An evidence-based review. Drug Safety, 33(9), 713–726. https://doi.org/10.2165/11536520-000000000-00000
Zhao, C. C., Zhai, Y. J., Hu, Z. J., Huo, Y., Li, Z. Q., & Zhu, G. J. (2022). Efficacy and safety of methylene blue in patients with vasodilatory shock: A systematic review and meta-analysis. Frontiers in Medicine (Lausanne), 9, 950596. https://doi.org/10.3389/fmed.2022.950596
Zheng, X., Ma, X., Li, T., Shi, W., & Zhang, Y. (2020). Effect of different drugs and drug combinations on killing stationary phase and biofilms recovered cells of Bartonella henselae in vitro. BMC Microbiology, 20(1), 87. https://doi.org/10.1186/s12866-020-01777-9
Zhou, L., Flores, J., Noël, A., Beauchet, O., Sjöström, P. J., & LeBlanc, A. C. (2019). Methylene blue inhibits Caspase-6 activity, and reverses Caspase-6-induced cognitive impairment and neuroinflammation in aged mice. Acta Neuropathologica Communications, 7(1), 210. https://doi.org/10.1186/s40478-019-0856-6
Zuschlag, Z. D., Warren, M. W., & Schultz, S. K. (2018). Serotonin toxicity and urinary analgesics: A case report and systematic literature review of methylene blue-induced serotonin syndrome. Psychosomatics, 59(6), 539–546. https://doi.org/10.1016/j.psym.2018.06.012